تنش رطوبتی در طول گرده افشانی ذرت (Zea maysL.) می تواند تا حد زیادی مقدار بذر را کاهش دهد، با این حال اطلاعات کمی در مورد اثرات وضعیت آبی گیاه بر رشد گل های نر و ماده موجود است. هدف از این مطالعه ایجاد تیمارهای مختلف تنش خشکی در طول گردهافشانی و اندازهگیری همزمانی رشد گلهای نر و ماده، زندهمانی گردهها و نرخ افزایش طول سیلک ها در روز بود. بذور هیبرید سینگل کراس در مزرعه و درون گلدان های مخصوص کشت شدند و در مرحله ظهور گل تاجی تحت تیمارهای مختلف خشکی قرار گرفتند. در زمان ظهور تاسل در مقایسه با گیاهان شاهد خوب آبیاری، تیمارهای خشکی خفیف (بدون پژمردگی قابل مشاهده) و شدید (پژمردگی قابل مشاهده) فاصله بین اولین سیلک ها تا اولین گرده ها را به ترتیب به طور متوسط 3 و 4 روز افزایش دادند. افزایش کمبود رطوبت باعث ایجاد تغییری در جوانه زنی گرده در شرایط آزمایشگاهی نشد، حتی زمانی که شدیدترین تیمار خشکسالی باعث علائم مشهود پژمردگی در میانه روز و پیری برگ های پایین بوته شد. اندازهگیریهای روزانه افزایش طول سیلک ها را نشان میدهد که در روزهای آفتابی، بیشترین افزایش رشد سیلک ها در شب اتفاق میافتد، زمانی که پتانسیل آب درون بلال در بیشترین میزان خود است. در پتانسیلهای آبی برگ مشابه در هنگام صبح، گیاهان تحت تنش نسبت به گیاهانی که به خوبی آبیاری شده بودند، میزان رشد طولی سیلک کمتری داشتند. رشد مثبت سیلک ها در پتانسیل های آبی بلال در حدود 9 بار در گیاهان تحت تنش و در 14 بار در گیاهان با آبیاری خوب متوقف شد، که نشان می دهد عواملی غیر از پتانسیل آب نیز ممکن است سرعت رشد سیلک را تنظیم و تحت تاثیر قرار دهند. نتیجهگیری میشود که خشکسالی با شروع گلدهی تأثیر بیشتری بر رشد گلهای ماده نسبت به نر دارد.
بالاترین عملکرد ذرت (Zea mays L.) در کمربند ذرت خیز ایالات متحده با بارندگی بالاتر از میانگین در طول ماه جولای(تیر و مرداد) و بارندگی معمول در طول سال به عمل آمده است است. (Thompson, 1969) حساس ترین بخش چرخه زندگی ذرت به خشکسالی ، رشد گل و لقاح می باشد که در ماه جولای رخ می دهد (سالتراند گود، 1967). دورههای کوتاه تنش خشکی زمانی منجر به بیشترین کاهش عملکرد میشود که تنش در طی ظهور سیلک و گردهافشانی اعمال شود (کلاسن و شاو، 1970). تنش خشکی در طول دوره طهور سیلک ها باعث ایجاد اختلال و عدم تشکیل مادگی دانه ها در بلال می شود و این کمبود و کچلی حاصل را نمی توان با بارش ( آبیاری) بهینه پس از این دوره تنش جبران کرد (رابینز و دومینگو، 1953). در مطالعات میدانی، تأخیر بین گرده افشانی و ظهور سیلک به عنوان یکی از دلایل اصلی کاهش عملکرد در نظر گرفته شده است. با این حال، تنش خشکی در طول گلدهی نیز در تأخیر رشد و تکامل دانه گرده دخیل است (سالتر و گود، 1967). ماس و داونی (1971)، در آزمایشهای گلخانهای، از تیمارهای خشکی در طول رشد اولیه کیسههای جنینی استفاده کردند و عقیمی مادگی را به دلیل سقط کیسه جنین مشاهده کردند. با این حال، هنکل (1964) پیشنهاد کرد که به طور کلی، گل نر نسبت به گل ماده به خشکی حساستر است.
در حالی که کاهش عملکرد ناشی از تنش خشکی به وضوح مشاهده شده است، عوامل فیزیولوژیکی که مسئول هستند کمترین میزان را در چرخه زندگی گیاه درک کرده اند (بویر و مک فرسون، 1975). در مزرعه، درجه حرارت بالا، رطوبت کم و رطوبت کم خاک اغلب به طور همزمان اتفاق میافتد و نسبت دادن خسارت به یک علت خاص را دشوار میکند (کمپ، 1954). هدف از این مطالعه تعیین واضحتر این بود که چه بخشهایی تحت تأثیر تنش خشکی رخداده در ظهور منگوله قرار میگیرند. با کاشت مزرعه در چندین تاریخ، آزمایشها طیف وسیعی از محیطها را در تلاش برای حذف دما بهعنوان یک متغیر اصلی نمونهبرداری کردند. عواملی که تحت رژیمهای تنش رطوبتی مختلف مورد تحقیق قرار گرفتند، شامل همگامسازی رشد گلهای نر و ماده، زندهمانی گرده و نرخهای ابریشمیشدگی روزانه بود.
مواد و روش ها
گیاهان مورد استفاده در این مطالعه در گلدان های حاوی تقریباً 18 کیلوگرم مخلوط خاک خشک رشد کردند. در یک قطعه مزرعه در مزرعه کشاورزی جنوبی، اوربانا، ایالت، گلدانها در ترانشههای کم عمق قرار گرفتند به طوری که گیاهان در عرض ردیف ۷۶ سانتیمتری با تراکم جمعیت ۵۰۰۰۰ بوته در هکتار قرار گرفتند. از ترانشه های کم عمق و گیاهان مرزی برای جلوگیری از تنش گرمایی به سیستم ریشه گلدانی استفاده شد. گیاهان مرزی همچنین به ایجاد رقابت بیشتر در میدان نرمال کمک کردند. گلدان ها با کیسه های پلاستیکی پوشانده شدند تا در صورت نیاز به استرس، کیسه ها را بتوان محکم در اطراف ساقه پایین پیچید تا از ورود بارندگی به گلدان جلوگیری شود. به این ترتیب، مقدار آب دریافتی توسط هر گیاه قابل تنظیم است. گیاهان در طول رشد رویشی به صورت بارور و آبیاری مناسب نگهداری میشوند. در زمان ظهور منگوله، رژیم های مختلف وضعیت آب آغاز شد. سه رژیم آبی مورد استفاده از نظر پتانسیل آب برگ گوش با قرائت در 1530 CDT اندازه گیری شدند. اینها عبارت بودند از: گیاهان شاهد در دمای 7 تا 11 بار، گیاهان تنش ملایم در دمای 11 تا 16 بار و گیاهان با کمبود شدید آب در دمای 16 تا 18 بار نگهداری شدند. پتانسیل آب با یک بمب فشار میدانی قابل حمل اندازه گیری شد (De Roo, 1969). پارچه پنیر مرطوب در پایین محفظه فشار برای کمک به کنترل رطوبت نگه داشته شد.
همگام سازی گرده افشانی
هدف از این آزمایش تعیین اینکه آیا همگام سازی اجزای تشکیل دهنده فرآیند گرده افشانی به دلیل وضعیت آب گیاه تغییر کرده است یا خیر. این آزمایش در سال 1978 بر روی هیبریدی “FR632 × FR619” انجام شد. از 12 تکرار فاکتوریل 6×3 در آرایش کرت های خرد شده از بلوک های کامل تصادفی استفاده شد. تاریخ کاشت به صورت کرتهای کامل و شامل شش کاشت متوالی در فواصل حدود 10 روزه بود که از 3 می 1978 شروع شد. سه تیمار تنش خشکی در تاریخ کاشت بهطور تصادفی انتخاب شدند. برای تعیین همزمانی، تاریخها برای جوانهزنی، سبز شدن پیشپروفیل، شروع جوانهزنی اولیه و شروع جوانهزنی اولیه ثبت شد. . هر یک از این مراحل رشد به شرح زیر تعریف شده است: (الف) ظهور منگوله تاریخ اولین اکستروژن منگوله از نقش برگ بود. (ب) ظهور پروفیل ظهور تاریخ از غلاف برگ گوش ساختار برگ مانند مرتبط با ساقه بلال بود. (ج) گلدهی تاریخی بود که اولین بساک از گلوم منگوله بیرون کشیده شد. و (د) ابریشم زمانی بود که اولین ابریشم ها از شاخساره گوش بیرون آمدند.
زنده ماندن گرده و وضعیت آب گیاه
این آزمایش به منظور تعیین اثر پتانسیل آب گیاه بر زنده مانی گرده انجام شد.
در سال 1978، از طرح بلوک های کامل تصادفی با شش تکرار و سه تیمار تنش خشکی استفاده شد. تکرارها در طول زمان با تاریخ کاشت های متوالی در فواصل تقریباً 10 روزه از 3 می (اواسط اردیبهشت)آغاز شد. در سال 1978 هیبریدی FR632 × FR619 به عنوان ماده آزمایشی استفاده شد. این آزمایش در سال 1979 با استفاده از سه تکرار و دو هیبرید، FR632 × FR619 و ‘Mo17 × B73 تکرار شد. در هر دو سال، تیمارهای خشکسالی در مرحله جوانه زنی آغاز شد. گرده با انتخاب شاخه های تاسل از هیبریدهایی که در سه سطح مختلف تنش پتانسیل آبی بودند جمع آوری شد. سطح تنش خشکی گیاه در روز قبل از جمع آوری گرده تعیین شد و همچنین در روز جمع آوریگرده ها چک و تایید شد. این کار اطمینان حاصل کرد که گیاه حداقل 24 ساعت قبل از تعیین زنده ماندن گرده در سطح تنش بوده است. پانیکول های تاسل ها در صبح ساعت 0600 جمع آوری شد و بساک هایی که در آن روز خارج شده بودند به عنوان منبع استخراج گرده استفاده شدند. جوانه زنی گرده در محیط آزمایشگاهی بر روی محیط کشت آگار-نوبل انجام شد. روش جوانه زنی کوکاند والدن (1965) با افزایش غلظت آگار از 0.7 به 1.5 درصد اصلاح شد. از دمای محیط و رطوبت نسبی استفاده شد و قبل از ثبت شمارش جوانه زنی حدود 2 ساعت اجازه داده شد تا جوانه زنی ادامه یابد. دانههای گرده جوانهزده دانههایی بودند که دارای لولههای کامل به اندازه ی حداقل قطر یک دانه گرده بودند (کوک و والدن، 1965). دانه های ترکیده دارای توده ای نامنظم از سیتوپلاسم و دانه های نشاسته بودند و گرده های جوانه زده پس از آن هیچ فعالیتی نشان ندادند. نتایج جوانه زنی به عنوان درصد از تعداد کل گزارش می شود
 رشد سیلک ها تحت پتانسیلهای مختلف آب
رشد سیلک ها تحت پتانسیلهای مختلف آب
آزمایش سوم شامل اندازهگیریهای روزانه طول سیلک بود. یک مبدل خطی برای ثبت پیوسته افزایش طول ابریشم استفاده شد. یک چرخش مبدل یک تغییر ولتاژ از 0 تا 300 اینچ ایجاد کرد. حداکثر خطا در ± 0.02 میلی متر محاسبه شد. دستگاه مشابهی توسط Hsiao و همکاران (1970) برای اندازهگیری رشد طولی برگهای تک لپهای سالم ساخته شد. یک سیستم قرقره به مبدل خطی متصل شد. سیستم قرقره شامل یک دیسک فلزی بود که یک پیچ بدون سر به آن ذوب شده بود. نخ دوخت پلی استر محکم روی قرقره بسته شده بود. نخ دور دیسک فلزی پیچیده شد و حاوی وزنه ای بود که در انتهای آن بسته شده بود. سر دیگر نخ به ابریشم ها متصل است. همانطور که ابریشم ها طویل می شدند، وقتی کشش از نوار متصل شده برداشته شد شل شدن نوار توسط وزنه شمارنده گرفته شد و باعث چرخش دیسک فلزی و خروجی ولتاژ شد. ابریشم ها با لاستیک سیلیکون و یک کاتالیزور اکتاتات قلع به نخ متصل شدند. در یک آزمایش مقدماتی، منطقه مریستمی در پایه ابریشم تحت تاثیر لاستیک سیلیکون قرار نگرفت، و نرخ ازدیاد طول سیلک معادل سیلک های تیمار شده با تونون_سیلیکون بود. به دلیل ماهیت ظریف سیلک ها برای اندازه گیری میانگین رشد، حدود 20 تا 30 سیلک برای هر چسب استفاده شد.
برای جلوگیری از حرکت ساقه و بلال، یک گیره متصل به یک ستون فولادی محکم روی بلال قرار داده شد. نکته مهم این واقعیت بود که گرده افشانی از طویل شدن ابریشم جلوگیری می کند، در حالی که جلوگیری از گرده افشانی اجازه می دهد تا رشد سیلک ها ادامه یابد (جونز و نیول، 1948). بنابراین جعبه ای ساخته شد تا نوک بلال را محصور کند، در حالی که باقیمانده گیاه در معرض شرایط میدان طبیعی قرار داشت. سیستم قرقره مبدل در داخل جعبه قرار داشت. دو سیستم ساخته شد، و نرخ افزایش طول سیلک همیشه به طور همزمان روی گیاهان تحت تنش و شاهد اندازهگیری میشد.
نتایج
بخش هایی از همگام سازی رشد گل های نر و ماده تحت تأثیر تنش خشکی ناشی از گلدهی قرار گرفت (جدول 1). از آنجایی که ظهور پیشگیرانه تا حد زیادی قبل از اعمال تیمارهای تنش خشکی در ظهور تاسل رخ داده است، ظهور پروفیل تحت تأثیر تیمارهای تنش خشکی قرار نگرفت. فاصله زمانی بین ظهور تاسل و گرده افشانی نیز تحت تأثیر تیمار تنش قرار نگرفت، اما با تأخیر در تاریخ کاشت، این فاصله به طور کلی کاهش یافت.
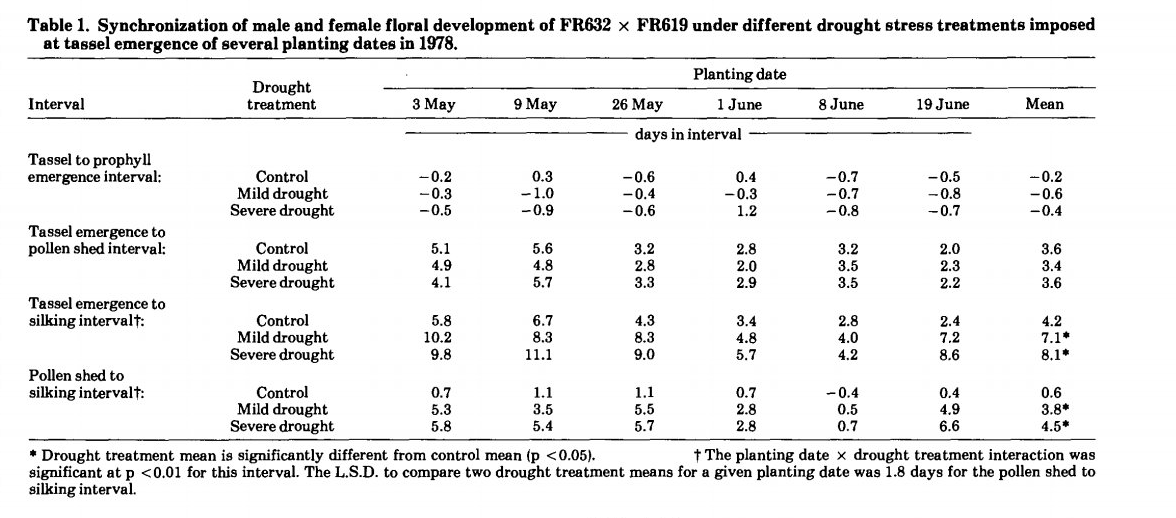
فاصله ظهور تاسل تا سیلک اولین مرحله رشدی مشاهده شده بود که توسط تیمارهای کمبود رطوبت افزایش یافت (جدول 1). این افزایش به دلیل افزایش تأخیر ظهور سیلک بود که با محاسبه فاصله گرده افشانی تا ظهور سیلک ها اندازهگیری شد. در مقایسه با گیاهان شاهد، تیمارهای تنش خفیف و شدید به ترتیب به طور متوسط 3 و 4 روز تاخیر ابریشم را افزایش دادند. تفاوت قابل توجهی در اثر متقابل تیمار خشکی و تاریخ کاشت برای تاخیر ظهور سیلک وجود داشت که عمدتاً به دلیل کاهش اثر کمبود رطوبت خاک در تاریخ کاشت 1 و 8 ژوئن(اواسط خرداد) بود. در طول دوره گرده افشانی بوته های روییده شده ازدر این تاریخ کاشت ها، اکثر روزها ابری و بارانی بود. کمبود رطوبت خاک ایجاد شد، اما تقاضای تبخیر و تعرق برای القای پتانسیلهای آب گیاهی مورد نظر در تیمارهای تنش خفیف و شدید کافی نبود. در طول دوره گرده افشانی گیاهان از تاریخ کاشت 19 ژوئن(اواخر خرداد)، هوای صاف بازگشته بود و کمبود رطوبت فزاینده دوباره باعث تاخیر شدید رشد سیلک ها شد.
با تأخیر در تاریخ کاشت، فاصله ظهور تاسل ها تا گرده افشانی و فاصله ظهور تاسل تا سیلک کاهش یافت. این اثر مستقل از ملاحظات کمبود آب بود. تغییرات در دوره نوری و/درجه روز رشد ممکن است کوتاه شدن این فواصل با کاشت تاخیری را توضیح دهد.
بر خلاف تاخیر در ظهور سیلک ها،افزایش کمبود رطوبت تنها اثرات جزئی و غیر قابل توجهی بر جوانه زنی گرده در شرایط آزمایشگاهی داشت (جدول 2).
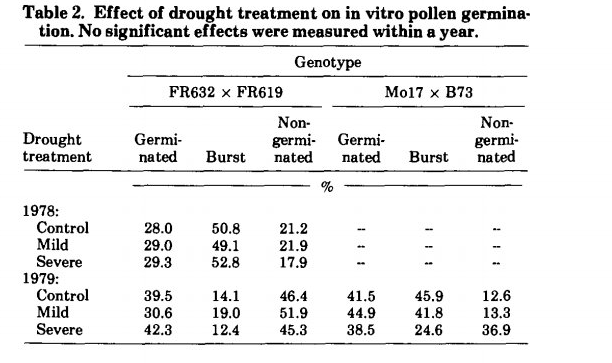
یک هیبرید تحت آزمایش و تیمار در سال 1978 و دوهیبرید آزمایش شده در سال 1979 در هر یک از سه تیمار تنش خشکی، درصد گرده های جوانه زده، ترکیده و جوانه نزده مشابهی داشتند. به نظر میرسد که FR632 × FR619 در سال 1979 درصد جوانهزنی گرده بیشتری نسبت به سال 1978 داشت، اما تحت تأثیر تیمار های تنش خشکی در هر دو سال قرار نگرفت. در طول هر دو سال، تیمار خشکی شدید باعث علائم آشکار پژمردگی و پیری برگهای زیرین شد. اثرات تکراری در سال 1978 قابل توجه بود اما نه در سال 1979. این موضوع تا حدودی قابل پذیرش است زیرا در سال 1978 تکرارها با اختلاف زمانی زیاد کاشته شدند، در حالی که در سال 1979 همه تکرارها در عرض 10 روز از یکدیگر کاشته شدند.
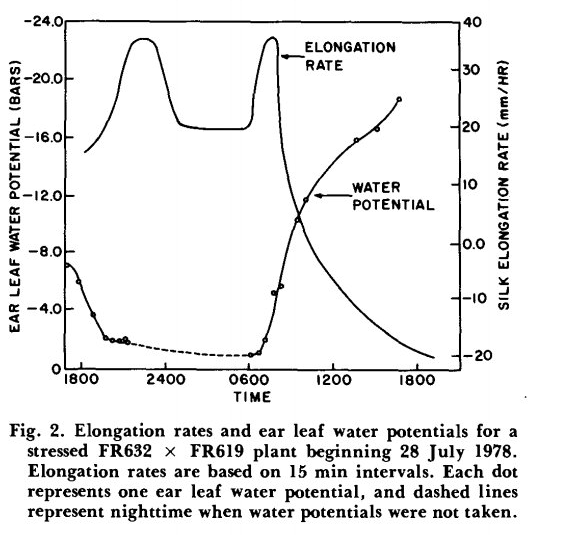
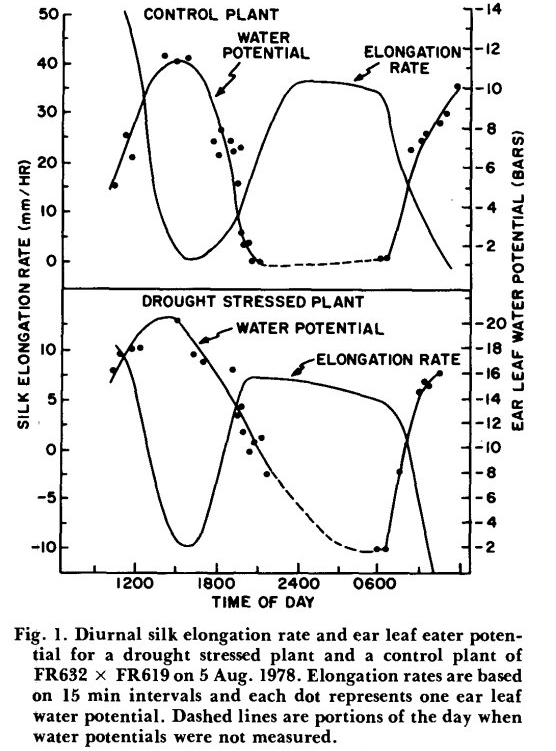
با تغییر در شرایط محیطی، هم نرخ افزایش طول سیلک ها و هم پتانسیل آب می تواند در مدت کوتاهی به شدت تغییر کند. این امر به ویژه برای گیاهان تحت تنش در اوایل صبح، زمانی که پتانسیل آب به سرعت کاهش مییابد، و برای گیاهانی که به خوبی آبیاری میشوند در غروب که پتانسیلهای آب به سرعت بهبود مییابند، صادق است. در ساعات شب، سرعت طویل شدن معمولاً از غروب خورشید تا نزدیک به ساعات 2400 یا 0100 در بیشترین میزان بود. ساعت ها پس از این زمان، افزایش طول سیلک کمی مانند شکل 1 یا به طرز چشمگیری مانند شکل 2 کاهش یافت. افزایش رشد طولی در اوایل صبح حدود یک ساعت پس از طلوع خورشید به وقوع پیوست. در انجام این آزمایش ها، فاکتور های مد نظر در یک گیاه تحت تنش خشکی همزمان با گیاه شاهد بدون تنش اندازه گیری شد. تفاوت در میزان رشد طولی شبانه سیلک ها در شکل 1 و 2 ظاهراً توسط عواملی غیر از تیمار خشکسالی ایجاد شده است، زیرا این تغییرات مستقل از تیمار تنش خشکی مشاهده شده است. هر نوع الگوی رشد طولی در شب، هم برای گیاهان تحت تنش هم برای گیاهان خوب آبیاری شده به دست آمده است. نرخ افزایش رشد سیلک ها از ساعات 8 صبح که اندازه گیری های روزانه در هر دو بوته های شاهد و تحت تیمارتنش خشکی جمع آوری شد، بر روی پتانسیل آب بلال کاهش یافت (شکل 3). مشاهدات به محدودهای از پتانسیلهای تقریباً برابر آب بلال برای بوته های شاهد و تحت تنش خشکی محدود شد. درپتانسیلهای مشابه آب بلال، گیاهان تحت تنش نسبت به گیاهانی که به خوبی آبیاری میشوند، نرخ افزایش طول سیلک کمتری داشتند. افزایش طول مثبت سیلکدر حدود 9- بار در گیاهان خشک شده و در 14- بار در گیاهان شاهد خوب آبیاری متوقف شد. در مقایسه با گیاهان شاهد، داده های پایین تر در گیاهان تحت تنش خشکی در پتانسیل های آبی بالاتر به دلیل کاهش سریع پتانسیل آبی برگ بلال در اوایل صبح با افزایش نیاز اتمسفری و تعرق است. اگرچه تیمار خشکی پراکندگی بیشتریاز نقاط داشت(انحراف از معیار در ظهور سیلک ها) ، اما شیب های مختلف دوخط نشان می دهد که کنترل رشد سیلک ها به تنهایی با پتانسیل آب در سال محدود نمی شود.
بحث
تنش خشکی در طول گلدهی باعث افزایش فاصله بین شروع گرده افشانی و ظهور سیلک ها شد. با استرس شدید، ظهور سیلک تا زمانی که تمام یا بیشتر گرده ریخته شد، به تعویق افتاد و در نتیجه تعداد زیادی بلال های بی ثمر یا ضعیف ایجاد شد. جنسن (1971) در آزمایش های نبراسکا تحت رژیم های آبی مختلف، همبستگی مثبت بالایی را بین تاخیر ظهور سیلک و عملکرد دانه گزارش کرد. در آزمایشهای تنش خشکی همچنین به این نتیجه رسیدیم که ظهور با تاخیر سیل و طویل شدن همزمان و بیش از حد، حساسترین بخش فرآیند هماهنگسازی گلها را نسبت به وضعیت آب گیاه نشان میدهد. در حالی که گاهی سیلک روی بوته های تحت تنش شدید خشکی ظاهر نمیشوند، سیلک ها در داخل پوشش برگ بلال باقی مانده و از گرده افشانی احتمالی جلوگیری میشود. عدم تأثیر تنش خشکی بر زندهمانی گردهها واکنشی غیرمنتظره بود. در مطالعه قبلی (Herreroand Johnson، 1981)، ما گزارش دادیم که گرده پذیری در شرایط آزمایشگاهی با قرار گرفتن تاسل ها در معرض تنش دمای بالا کاهش می یابد. جونز و نیوول (1948) بیشترین طول عمر را در زنده ماندن گرده های ریخته شده اگر گرده در شرایط دمای پایین و رطوبت نسبی بالا ذخیره شود گزارش کردند. نتایج ما حاکی از آن است که در طول نمو دانه گرده گرده، دمای بالا مضرتر از تنش خشکی است. هنکل (1964) تاکید کرد که مقاومت در برابر تنش دمای بالا همیشه با مقاومت به خشکی مرتبط نیست. مقاومت به خشکی گرده به دلیل وجود واکوئل کوچک درون دانه های گرده است (لینسکنز، 1964). اگر واکوئل سلول بزرگ باشد و تونوپلاست آن ظریف باشد تخریب سلول آسانتر می شود و مقاومت آن کاهش می یابد (ایلجین، 1957). توضیح احتمالی دیگر برای عدم اثر تنش خشکی بر جوانه زنی گرده ممکن است به دلیل بهبودمحتوای آب شبانه باشد که توسط آن تجربه شده است. گیاهان کشت شده در مزرعه حتی گیاهانی که در ساعات روز به شدت تحت تنش قرار میگرفتند، در طول شب به پتانسیلهای آبی بالا بازمیگردند. علاوه بر این، گرده افشانی در اوایل صبح اتفاق افتاد، قبل از اینکه پتانسیل آب بسیار کاهش یابد، که ممکن است به گرده کمک کند تا از اثرات منفی پتانسیل آبی کم گیاه در روز بگریزد.
 وضعیت روزانه افزایش طول سیلک ها به این دلیل مورد نظر است که مواد استیلار مورد نیاز برای ساخت لوله دانه ی گرده را فراهم می کند. لوله های گرده بین سلول های غلاف سیتوپلاسمی متراکم که آوند سیلک را احاطه کرده اند به سمت پایین حرکت می کند. همانطور که در شکل 1 و 2 می توان اشاره کرد، پتانسیل آب برگ بلال در معرض تغییرات روزانه است، با حداقل پتانسیل آب نزدیک به نیمه بعد از ظهر. این واقعیت که افزایش طول سیلک ها تقریبا در نیمه بعد از ظهر به کمترین میزان رشد طولی خود می رسند، نشان می دهد که طویل شدن سیلک ها تحت تاثیر وضعیت آب گیاه، عمدتاً در شب اتفاق میافتد و گردهها در صبح زود ریخته میشد، بنابراین با ابریشمهایی مواجه میشد که از شب قبل هنوز چروکیده بودند. در بخش عمده ای از کمربند ذرت ایالات متحده، شبنم اغلب در شب روی برگ ها جمع می شود، و حتی گیاهان تحت تنش شدید در بخش هایی از ساعات شب، شادابی را دوباره به دست می آورند.
وضعیت روزانه افزایش طول سیلک ها به این دلیل مورد نظر است که مواد استیلار مورد نیاز برای ساخت لوله دانه ی گرده را فراهم می کند. لوله های گرده بین سلول های غلاف سیتوپلاسمی متراکم که آوند سیلک را احاطه کرده اند به سمت پایین حرکت می کند. همانطور که در شکل 1 و 2 می توان اشاره کرد، پتانسیل آب برگ بلال در معرض تغییرات روزانه است، با حداقل پتانسیل آب نزدیک به نیمه بعد از ظهر. این واقعیت که افزایش طول سیلک ها تقریبا در نیمه بعد از ظهر به کمترین میزان رشد طولی خود می رسند، نشان می دهد که طویل شدن سیلک ها تحت تاثیر وضعیت آب گیاه، عمدتاً در شب اتفاق میافتد و گردهها در صبح زود ریخته میشد، بنابراین با ابریشمهایی مواجه میشد که از شب قبل هنوز چروکیده بودند. در بخش عمده ای از کمربند ذرت ایالات متحده، شبنم اغلب در شب روی برگ ها جمع می شود، و حتی گیاهان تحت تنش شدید در بخش هایی از ساعات شب، شادابی را دوباره به دست می آورند.
در آزمایشات اتاق رشد بویر (1970) گزارش داد که طویل شدن برگ ذرت تا حد زیادی در 4- بار مهار شد و در پتانسیل آب برگ نزدیک 8- بار به صفر رسید. همانطور که در شکل 3 نشان داده شده است، افزایش رشد طولی در سیلک ذرت به ترتیب در پتانسیل آب برگ بلال در حدود 9- و در گیاهان تحت تنش خشکی شده و14- بار آبیاری خوب متوقف شده است. بنابراین مانند برگهای ذرت، سرعت افزایش طول سیلک به شدت به وضعیت آب گیاه پاسخ میدهد. با این حال، پتانسیل آبی برگ بلال مشابه طویل شدن ابریشم در شرایط تنش خشکی، نسبت به گیاهانی که به خوبی آبیاری شده اند، نزولی می شود. همچنین به نظر می رسد که الگوهای طویل شدن سیلک در شب مستقل از وضعیت آب است (شکل 1 در مقابل شکل 2). اندازه گیری پتانسیل آب نقطه رشد سیلک ها به خوبی توسط پوشش های بلال محافظت می شود و بنابراین در معرض مستقیم کمتری برای نیاز اتمسفری و تعرق قرار می گیرد. این ممکن است باعث شود ابریشم در پتانسیل آب بالاتری نسبت به برگ گوش قرار گیرد. در برگهای ذرت، جایی که پتانسیلهای طولی و آب در سراسر برگ به طور همزمان اندازهگیری شده است، اندازهگیری اختلاف پتانسیل آب تا 3 بار از نقطه رشد تا نوک برگ غیرمعمول نیست (J. S. Boyer)نتیجهگیری میشود که در مزرعه پتانسیلهای آبی کم گیاه ناشی از کمبود آب در خاک، در مقایسه با تأثیرات روی زندهمانی گرده، تأثیر بسیار بیشتری بر ازدیاد طول ابریشم و هماهنگسازی متعاقب آن گرده افشانی و ظهور سیلک دارد. گردهزایی در شرایط آزمایشگاهی تحتتاثیر پتانسیلهای آبی گیاه قرار نگرفت تا حدی که پژمردگی قابل مشاهده و پیری برگهای پایین را ایجاد کند. تحت شرایط کمبود رطوبت خاک در مزرعه، پتانسیلهای آب گیاه در ساعات شب بهبود مییابند، و اجازه میدهد که اکثر ازدیاد طول ابریشم در این زمان رخ دهد. بازیابی رطوبتی شبانه همچنین ممکن است اثر محدود تنش خشکی بر زنده ماندن گرده ها را جبران کند.




